di Carlo Matessi
Introduzione
Nella fascia di media collina dell'Oltrepò Pavese esiste una popolazione di Zerynthia sp. individuata e tenuta in osservazione dal nostro socio Eugenio Balestrazzi da circa venti anni. Poiché il numero di adulti avvistati in volo di anno in anno è apparso sempre esiguo e l'area piuttosto ristretta occupata dalla popolazione è circondata da appezzamenti soggetti ad intensa attività agricola, considerata la grande importanza faunistica della specie si è ritenuto opportuno avviare un'indagine demografica ed ecologica volta a stimare nel modo più accurato possibile la dimensione della popolazione assieme ad altri parametri come: il periodo di volo, la condizione fisica degli adulti, il rapporto sessi, i tassi di reclutamento/immigrazione e di mortalità/emigrazione, la disponibilità di risorse alimentari per le larve (Aristolochia rotunda), la quantità totale di uova deposte, etc. Inoltre, da quando è stato dimostrato (Dapporto, 2009) che in Italia esistono due distinte specie di Zerynthia, Z. polyxena e la più meridionale Z. cassandra, ipoteticamente separate geograficamente dal corso del fiume Po, era urgente stabilire a quale delle due specie la nostra popolazione appartenesse. Pensiamo che la disponibilità di attendibili informazioni di questo genere sia indispensabile per orientare l'elaborazione di un eventuale futuro piano di salvaguardia della popolazione. La prima iniziativa in questa direzione è stata attuata nella primavera del 2018.
L'area di studio
L'area oggetto della nostra indagine, Sito Zerinzie (SZ), è un incolto di circa 5.7 ettari di superficie, disposto su un leggero declivio rivolto ad ovest, tra le quote di 500 m e 560 m slm, delimitato da un bosco di latifoglie ad est, vigneti e seminativi sui restanti lati. L'area è utilizzata stagionalmente per il pascolo di bovini. Abbiamo individuato una suddivisione del sito in 6 settori contigui (A, B, ...,F) diversi per superficie e distinguibili sia per le caratteristiche del suolo e della vegetazione che per la presenza di limiti naturali.L'area oggetto della nostra indagine, Sito Zerinzie (SZ), è un incolto di circa 5.7 ettari di superficie, disposto su un leggero declivio rivolto ad ovest, tra le quote di 500 m e 560 m slm, delimitato da un bosco di latifoglie ad est, vigneti e seminativi sui restanti lati. L'area è utilizzata stagionalmente per il pascolo di bovini. Abbiamo individuato una suddivisione del sito in 6 settori contigui (A, B, ...,F) diversi per superficie e distinguibili sia per le caratteristiche del suolo e della vegetazione che per la presenza di limiti naturali.
In tutti i settori Aristolochia rotunda è abbondante ma la distribuzione delle pianticelle è a macchie. In un piccolo studio pilota sul SZ da me fatto nella primavera del 2016, tra 22 quadrati di 1 metro di lato, scelti in modo rigorosamente casuale, solo 5 (23%) contenevano almeno una pianticella, mentre in altri 15 quadrati casuali contenenti aristolochia vi erano in media 11.6 pianticelle per quadrato. Volendo estrapolare questi dati molto approssimativi si potrebbe concludere che nel 2016 la disponibilità complessiva di aristolochia nel SZ è stimabile in 150273 pianticelle, con un coefficiente di variazione (rapporto tra l'errore standard ed il valore della stima) del 43%.In tutti i settori Aristolochia rotunda è abbondante ma la distribuzione delle pianticelle è a macchie. In un piccolo studio pilota sul SZ da me fatto nella primavera del 2016, tra 22 quadrati di 1 metro di lato, scelti in modo rigorosamente casuale, solo 5 (23%) contenevano almeno una pianticella, mentre in altri 15 quadrati casuali contenenti aristolochia vi erano in media 11.6 pianticelle per quadrato. Volendo estrapolare questi dati molto approssimativi si potrebbe concludere che nel 2016 la disponibilità complessiva di aristolochia nel SZ è stimabile in 150273 pianticelle, con un coefficiente di variazione (rapporto tra l'errore standard ed il valore della stima) del 43%.
Svolgimento dell'indagine 2018
In preparazione dell'indagine vera e propria il SZ è stato ispezionato in tre occasioni, il 7, il 17 ed il 21 aprile, per accertare la presenza delle prime pianticelle di aristolochia e dei primi adulti di Zerinzia e solo il 21 si sono viste alcune giovani pianticelle (12-25 cm), alcune con fiore, e 4 adulti in volo tra cui tre maschi.
Nel calendario inizialmente stabilito per lo studio erano previste 3-4 serie di uscite sul campo, di 4 giorni consecutivi ognuna, da eseguirsi in settimane consecutive cominciando da lunedì 23 aprile. Tuttavia il sopravvenire di maltempo ci ha costretto ad annullare 3 uscite nella seconda settimana e ad alterare la successione di uscite nella terza settimana, come mostrato nella Tabella 1. Inoltre, avendo constatato una rapida diminuzione delle presenze di zerinzie, alla fine della terza settimana abbiamo deciso di terminare lo studio.
TABELLA 1
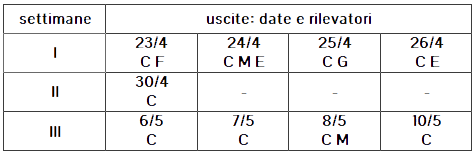
Le sigle indicano i rilevatori attivi ad ogni uscita:
C=Carlo Matessi, E=Eugenio Balestrazzi, F=Francesco Gatti, G=Gianluca Ancarani, M=Michela Villa
Generalmente i rilevamenti iniziavano tra le 9:30 e le 11:00, in funzione del tempo previsto nella giornata, e si concludevano tra le 14:00 e le 17:30, secondo la presenza di adulti e l'evoluzione delle condizioni metereologiche. Come indicato dalla tabella, complessivamente 5 rilevatori hanno partecipato allo studio, con presenze diverse nei diversi giorni. Le ore-persona dedicate in totale allo studio sono state 61.
Tutte le persone che hanno partecipato alle attività di campo, incluso l'autore di questa relazione, sono stati autorizzati a manipolare esemplari di Zerynthia sp. con atto del Ministero dell'Ambiente del 13/03/2018 no. 005061.
Procedura della raccolta dati
Per la raccolta dei dati necessari alla stima della dimensione della popolazione abbiamo adottato il metodo di "Cattura-Marcatura-Ricattura" (CMR). In ogni giornata i diversi settori del SZ sono visitati in successione, attraversando ciascuno ripetutamente in lungo e in largo in cerca di adulti di zerinzie in volo, tentandone la cattura ad ogni avvistamento. Per ciascun esemplare catturato ed estratto delicatamente dal retino sono immediatamente eseguite le seguenti operazioni: (i) si registra l'ora ed il settore di cattura; (ii) si identifica il sesso ispezionando l'estremità addominale; (iii) si misura l'ampiezza dell'ala anteriore sinistra tra la radice di attacco al torace e l'apice; (v) se l'esemplare non era stato mai catturato in precedenti uscite viene marcato sull'ala sinistra e se necessario anche sull'ala destra, con pennarello rosso permanente morbido, secondo un codice a punti per la numerazione individuale progressiva concepito da P. R. Ehrlich (Ehrlich e Davidson, 1960); (vi) se invece l'esemplare risulta già marcato viene soltanto identificato e registrato il suo numero e ogni altra ispezione e misurazione è naturalmente evitata; (vii) l'esemplare esaminato è riposto ad ali chiuse in una bustina per farfalle sulla quale sono stati già registrati tutti i dati sopra elencati e conservato in una scatola conservata al buio; (viii) i dati rilevati sono riportati anche nel taccuino delle note di campo; (ix) al termine della giornata gli esemplari catturati e riposti nelle bustine sono rilasciati, ciascuno approssimativamente al centro dello stesso settore in cui era stato catturato in giornata.
La manipolazione degli esemplari era eseguita da una sola persona mentre un secondo operatore registrava i dati rilevati sulla bustina che avrebbe ospitato l'esemplare catturato; nelle quattro uscite fatte da un unico rilevatore i dati erano trascritti sulla bustina subito dopo che vi era stato riposto l'esemplare esaminato. Ciascun esemplare rilasciato è sempre apparso in ottime condizioni e la vivacità con cui riprendeva il volo ne testimoniava l'ottima condizione fisica.
Le norme stabilite per il monitoraggio di specie protette (Bonelli et. al, 2016) richiedono per Zerynthia il rilascio immediato degli esemplari catturati. Questa prescrizione è indubbiamente pratica e raccomandabile nel caso in cui l'indagine si esaurisca in due sole sessioni, dedicate rispettivamente alla cattura-marcatura e alla ricattura e non sia necessario contrassegnare gli esemplari con numeri distinti. Nel nostro caso, in cui per le ragioni più sotto esposte è stato assolutamente indispensabile effettuare ripetute sessioni in giorni e settimane successive, abbiamo valutato che si dovesse agire in difformità da questa norma, adottando invece la procedura sopra indicata ai punti (vii) e (ix), proposta e utilizzata da autorità indiscusse nel campo dell'ecologia dei lepidotteri (Ehrlich e Davidson, 1960; Ehrlich, 1965; Ehrlich and Mason, 1966; Hellmann et al, 2004). Nella nostra situazione ogni volta che un esemplare già marcato viene catturato è necessario leggere dalle sue ali e registare il numero che lo identifica, operazione che può comportare una certa misura di stress per l'esemplare. Se gli esemplari marcati fossero rilasciati immediatamente, considerando la limitata mobilità della specie potrebbero essere ricatturati anche più volte nella stessa giornata, forse anche dopo brevi intervalli di tempo, con il rischio che lo stress da loro subito diventi eccessivo. Inoltre, poiché la ricattura di un esemplare già catturato nella medesima giornata deve essere ignorata e vanno registrate solo le nuove catture-marcature o le prime ricatture in giornata di esemplari marcati nei giorni precedenti, dato che pochi minuti o poche ore non garantiscono un adeguato rimescolamento della popolazione come richiesto dal metodo statistico CMR, fatta la lettura del numero marcato sull'esemplare sarebbe necessario verificare che esso non sia già stato registrato tra le catture del medesimo giorno. Perciò si può ben vedere che in tal modo sarebbe sottratto tempo alla raccolta utile di dati freschi e verrebbero amplificati i rischi di errore nella registrazione dei dati ogni qual volta venisse ripreso un esemplare già visto in giornata.
Esposizione sommaria dei risultati
In questa sede non saranno esposti i risultati di una rigorosa analisi statistica e valutazione critica delle informazioni raccolte, che non sono ancora state completate per la loro natura tutt'altro che elementare dovuta alle caratteristiche specifiche della nostra indagine, ma che saranno oggetto di una futura relazione scientifica. Mi limito pertanto ad una sommaria esposizione dei dati grezzi da noi raccolti allo scopo di mostrare la quantità e la qualità del lavoro svolto, insieme alla particolare difficoltà ad ottenere dati molto accurati in una popolazione esigua come la nostra e con i limitati mezzi di cui una semplice associazione di volontari può disporre.
Prima di iniziare la parte sistematica dello studio ci siamo preoccupati di identificare la specie di Zerynthia cui appartiene la nostra popolazione. Ciò si può fare semplicemente e rapidamente con un esame non cruento dei genitali maschili (valvae), che sono facilmente visibili dato che il maschio, trattenuto in posizione supina ed eventualmente solleticato gentilmente sull'addome, estrude completamente il perifallo mettendone in chiara evidenza le valvae. In questo modo, in base alla diagnosi fornita da Dapporto (2009) abbiamo verificato che la nostra popolazione appartiene alla specie Z. cassandra. La prima evidenza certa, documentata da foto, di cui si mostra un esempio nella Figura 1, e da un unico esemplare maschio sacrificato e conservato, è stata poi frequentemente confermata in maschi catturati durante la parte sistematica dell'indagine.
Negli anni precedenti un attendibile indizio del risultato ora provato dai genitali maschili ci era stato fornito dall'osservazione in SZ di bruchi di Zerynthia.
FIGURA 1

Fotografie delle valvae di maschi di Zerynthia. Da sinistra a destra: maschio della popolazione del SZ; preparato di genitali di maschio di Z. cassandra; preparato di genitali di maschio di Z. polyxena. La prima foto è del nostro socio G. Ancarani; le due foto di preparati sono tratte da Dapporto (2009). Nella prima foto è evidente l'apofisi appuntita in cui si prolunga il margine superiore delle valvae, caratteristica di Cassandra ed assente in Polissena.
FIGURA 2

Foto di larva di Zerynthia scattata nel 2016 da G. Ancarani al SZ
Dalle informazioni riferiteci da diversi colleghi e dalle mie stesse osservazioni su altre popolazioni di Z. cassandra si ricava la convinzione che le larve di Cassandra abbiano un carattere che le differenzia da quelle di Polissena. Gli scoli che adornano in 4 file dorsali e laterali le larve in Cassandra sono interamente di colore arancio (Figura 2) mentre terminano in una punta di colore nero in Polissena.
La Tabella 2 mostra alcuni dati riassuntivi della nostra ricerca. Considerando che i totali mostrati riguardano 9 giorni, la media di catture e ricatture giornaliere è rispettivamente di 8.3 e 2.3 individui. Come si vede il numero di individui giornalmente avvistabili nel SZ è sempre molto piccolo. Una stima delle dimensioni della popolazione (adulti presenti in ogni dato giorno) basata sui dati di singole giornate è facilmente calcolabile ma per l'esiguità dei dati sarebbe affetta da un'incertezza statistica molto elevata, anche se già si capisce che comunque in totale non può discostarsi troppo da 50-60 individui. È questa la ragione per cui si è stabilito di ripetere l'indagine in più giornate, per migliorare la qualità delle stime con un maggior numero di dati. In questo modo però si deve tener conto del fatto che mortalità/emigrazione e reclutamento/immigrazione hanno la possibilità di modificare la popolazione e diventa perciò necessario stimare in modo appropriato anche l'effetto di questi eventi, mediante analisi statistiche sostanzialmente più complesse. Nel caso presente la difficoltà è aggravata dalla mancanza di catture e marcature nella seconda settimana, a parte il primo giorno (vedi Tab. 1). Il confronto di questi dati con quelli della prima settimana e di quelli della terza con questi avrebbe potuto mettere in evidenza eventuali cambiamenti subiti dalla popolazione dopo un intervallo di 7-10 giorni, un arco temporale particolarmente significativo visto che il più lungo intervallo che abbiamo osservato tra cattura e ricattura di un individuo è stato di 11 giorni, una prima seppur grezza indicazione sulla permanenza in situ degli adulti.
TABELLA 2
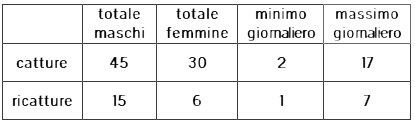
I massimi e minimi giornalieri comprendono maschi e femmine. I massimi sono stati realizzati il terzo giorno di indagine quando le condizioni metereologiche erano ottime. I minimi si sono verificati nell'ultimo giorno quando chiaramente la popolazione era in esaurimento e le condizioni metereologiche erano mediocri e seguivano giornate di mal tempo.
La Tabella 2 mostra che le catture di maschi eccedono del 50% quelle di femmine, una deviazione significativa dall'attesa parità numerica tra i sessi. Piuttosto che ad una reale deviazione da un rapporto sessi di 1:1 alla nascita, la deviazione osservata sarà più probabilmente dovuta alla maggiore mobilità dei maschi, comune tra i Ropaloceri, o anche ad un leggero ritardo nello sfarfallamento delle femmine. Nell'ultima settima si sono fatte 8 catture di femmine e 6 di maschi.
Mentre i settori A, B, D ed E del SZ sono stati perlustrati con pari intensità nella nostra indagine, i settori C ed F sono stati abbastanza trascurati a causa della maggiore difficoltà a percorrerli per la pendenza relativamente pronunciata. Di conseguenza, sebbene essi coprano il 25% della superficie totale di SZ, vi sono state fatte soltanto 3 catture (4% del totale). Escludendo questi settori, nella Tabella 3 sono mostrate le quote relative di superficie e catture che competono agli altri 4 settori. Complessivamente le catture sono abbastanza proporzionate alle superfici, anche se il settore D appare relativamente più produttivo ed il settore E meno produttivo. In mancanza di dati appropriati non è possibile verificare se questa differenza possa essere attribuita ad una disparità nell'abbondanza di aristolochia tra i due settori.
TABELLA 3

Per i settori di SZ più visitati si mostra in % l'ammontare relativo della superficie occupata e delle catture ivi effettuate
Indicazioni per studi futuri
È evidente la necessità di ripetere, se possibile con maggior sforzo, questa indagine il prossimo anno, sperando che la situazione metereologica, causa principale dell'insufficienza dei dati attuali, sia più favorevole. Un possibile rimedio alla scarsità di dati potrebbe venire dall'effettuare le uscite sul campo ogni giorno metereologicamente favorevole, evitando le interruzioni settimanali programmate questa prima volta. Con l'esperienza acquisita si è visto che le zerinzie, diversamente da altre specie non disdegnano di volare fino ad ore tarde prossime al crepuscolo. Quindi anche un prolungamento della permanenza giornaliera sul campo potrebbe aiutare. Inoltre ci si deve sforzare di perlustrare più intensamente i settori trascurati C ed F che sicuramente sono visitati dalle zerinzie ed ospitano aristolochia.
Lo studio della popolazione adulta dovrebbe essere utilmente integrato da un'indagine sulla copertura della superficie di SZ da parte di Aristolochia rotunda secondo il metodo (quadrati casuali) adottato nella mia esperienza pilota del 2016; ma il numero di quadrati dispiegati dovrebbe essere portato almeno fino a 100 per ridurre da 43% almeno fino a 20% circa il coefficiente di variazione della stima del numero totale di pianticelle. Inoltre i sondaggi dovrebbero coinvolgere anche i settori C, E, F ignorati nel 2016.
Per una esauriente analisi della popolazione di Cassandra l'attenzione dovrebbe essere rivolta oltre che agli adulti anche alle uova, ma possibilmente anche alle larve. In questi casi le analisi statistiche richieste sono molto più semplici ma lo sforzo fisico degli operatori è molto maggiore perché i conteggi dovrebbero essere fatti sulle singole pianticelle di aristolochia rinvenute all'interno di quadrati casuali. Per quanto riguarda le uova, molto facili ad individuare (Figura 3), il censimento può essere fatto contestualmente al censimento della pianta, limitandolo ad un piccolo numero fisso di pianticelle presenti in un quadrato.
Questo è ciò che ho fatto nella mia indagine pilota del 2016. In ognuno dei 15 quadrati contenenti piantine di Aristolochia ho esaminato 5 piantine, o tutte quelle presenti se in minor numero, contando le uova di Zerinzia che vi si trovassero. Delle 72 piantine esaminate 65 (90%) erano prive di uova, mentre
FIGURA 3

Quattro uova di Z. cassandra (bianche) e una ninfa di afide (gialla) sulla pagine inferiore di una foglia di Aristolochia rotunda al SZ
tra le altre 7 piantine ho trovato in media 2 uova per piantina. Volendo azzardarci ad estrapolare questi pochi dati a tutta l'area del SZ, dove abbiamo crudamente stimato il numero totale di piante essere pari a 150273 (vedi sopra), si potrebbe concludere che nel 2016 la popolazione totale di uova di Zerynthia cassandra ammontava a circa 30000 unità, stima che però ha un coefficiente di variazione del 64% ed inoltre, pur riferendosi all'intera superficie del SZ si basa soltanto su dati rilevati nei soli settori, A, B e D, ispezionati nel 2016. Se questi calcoli fossero attendibili potremmo concludere che la fertilità delle femmine di Cassandra nel SZ è molto elevata (dell'ordine di 500-1000 uova per femmina) ma la sopravvivenza fino allo stadio adulto delle loro uova è estremamente bassa (dell'ordine del 2-3‰). Una valutazione della popolazione di larve potrebbe essere condotta in modo simile ma potrebbe essere di più difficile e problematica esecuzione perché, dovendo avvenire necessariamente a stagione più avanzata (fine maggio-primi di giugno), la copertura erbosa generale divenuta molto più fitta tenderà ad ostacolare l'individuazione e l'ispezione delle pianticelle di Aristolochia, con la conseguente maggiore probabilità di danni alle pianticelle e di disturbo alle larve, che potrebbero sottrarsi alla vista o addirittura subire esse stesse qualche danno. Nel corso del 2018 sono state individuate nell'Oltrepò Pavese altre due popolazioni di Zerynthia sp. in località piuttosto lontane tra loro e da quella del SZ (circa 20-25 km), accertate per la presenza di uova su Aristolochia rotunda e di adulti, tra cui una femmina che deponeva su A. rotunda, di cui ignoriamo del tutto la consistenza. È evidente che indagini del tutto analoghe a quelle effettuate o prospettate per il SZ dovranno essere rivolte anche a queste nuove popolazioni, ammesso che sia possibile reclutare le forze necessarie a questo compito.
Bibliografia
Scarica PDF